New World Primates Nutrition
Barrueta-Acevedo F.M.
Introducción
Los primates del nuevo mundo o platirrinos, son un grupo conformado por cinco familias nativas de Centro y Sudamérica: Cebidae, Aotidae, Pitheciidae, Atelidae y Callitrichidae. Dentro de los cébidos, encontramos a los monos carablanca y los monos ardilla. Se denomina Aotus a un grupo de primates nocturnos distribuidos en los bosques tropicales de Centro y Sudamérica. Los pitécidos incluyen a los monos saki y uakaris, pertenecientes a los géneros Pithecia, Chiropotes, Cacajao y Callicebus. Los atélidos, anteriormente incluidos en la familia Cebidae, se componen por los monos araña, congo y lanudos. Finalmente, los calitrícidos, para algunos aún considerados dentro de los cébidos, están representados por las marmosetas y los tamarinos, e incluyen los géneros Cebuella, Callibella, Mico, Callithrix, Callimico, Saguinus y Leontopithecus (Richard 2004, Chatterjee et al 2009).
Existen numerosos factores que llevan a la tenencia de primates en cautiverio, enfocados principalmente en la conservación de las especies. Para el caso de los primates del nuevo mundo, destacan los decomisos debidos al tráfico ilegal de fauna silvestre que llegan a centros de rescate o colecciones públicas o privadas, la tenencia en zoológicos con fines educativos y los centros de investigación biomédica.
La nutrición constituye una de las medidas más importantes de manejo para el éxito en la supervivencia y reproducción de las especies, por lo que debe considerarse siempre que se esté trabajando con animales cautivos, considerando los hábitos alimenticios propios de cada especie, para evitar la presentación de enfermedades debidas a desbalances nutricionales. El estado nutricional de un animal está directamente relacionado con su alimentación. Un manejo nutricional correcto permite que el animal alcance todo su potencial en fases como el crecimiento, la reproducción y la lactancia, además de permitir una mayor longevidad. Asimismo, una alimentación adecuada permite al animal presentar respuestas más favorables a factores de estrés ambiental o a la presencia de patógenos.
Este trabajo pretende proporcionar al lector la información básica sobre la nutrición de primates del nuevo mundo en condiciones de cautiverio, a través del conocimiento general de la anatomía y fisiología del tracto digestivo de los mismos, las adaptaciones propias y los nutrientes indispensables para cada una de las familias que integran este grupo. Se incluye además, una descripción de las principales patologías nutricionales y algunas técnicas de alimentación que pueden ayudar a un mejor consumo de la dieta y al bienestar de los animales, dependiendo de sus condiciones individuales.
Hábitos alimenticios
Los hábitos alimenticios de los primates del nuevo mundo en vida libre, son muy variados e incluyen diferentes alimentos, tales como frutas, hojas, semillas, flores, invertebrados y hasta pequeños mamíferos y reptiles, por mencionar algunos. Cabe señalar que cada especie tiene diferentes preferencias, por lo que a continuación se especifican los alimentos consumidos con mayor frecuencia por cada una de las familias que integran el grupo de los platirrinos.
Cebidae
Consumen principalmente frutos, semillas, flores, peciolos, hojas tiernas, larvas de insectos, huevos de aves y pequeños invertebrados. Los monos carablanca incluso llegan a consumir vertebrados. Los monos ardilla tienden a ser más insectívoros durante la escasez de frutas (Richard 2004).
Aotidae
Esta familia de primates nocturnos consumen una amplia variedad de alimentos, dentro de los que se incluyen frutos, néctar, hojas, insectos, lagartijas, aves y pequeños mamíferos (Umaña 1984).
Pitheciidae
Dentro de esta familia se encuentra el género Callicebus, que se alimenta principalmente de frutas, hojas, invertebrados y flores (Barbosa 2008). Chiropotes y Pithecia consumen principalmente semillas, pulpa de frutos maduros, hojas e insectos (Riveros & Ferreira 2001, Pinto 2008).
Atelidae
En esta familia se encuentra el género Ateles, que se alimenta principalmente de frutos, flores, hojas tiernas y pequeños vertebrados. También tenemos al género Alouatta, probablemente el único primate vegetariano estricto, que se alimenta básicamente de hojas, frutos, flores, brotes y yemas. Por último, el género Lagothrix, que también forma parte de esta familia, se alimenta principalmente de frutos, pero también consume flores, semillas, hojas tiernas y pequeños invertebrados (Pastor-Nieto 2004, Richard 2004).
Callitrichidae.
Los platirrinos pertenecientes a esta familia son, como la mayor parte de los primates, omnívoros, y su dieta se basa en exudados de árboles, insectos y, ocasionalmente, frutos, brotes y néctar (Mayor & López 2012). Se ha reportado que en la naturaleza, los calitrícidos de mayor tamaño tienden a consumir insectos y frutos, mientras que los de menor tamaño consumen insectos y exudados, como gomas, savia y látex (Crissey et al. 2003).
Anatomía del aparato digestivo
Generalidades
El sistema digestivo de los platirrinos es corto y simplificado (Varela 2005). Debido a las variaciones entre especies, el sistema digestivo de los vertebrados se describe de mejor manera mediante su división por regiones, comenzando en los componentes bucales, esófago, estómago, intestino delgado e intestino grueso. De gran importancia son también sus órganos adicionales, incluyendo las glándulas salivales, páncreas y el sistema hepatobiliar (Finegan & Stevens 2000).
El tracto digestivo tiende a ser más corto y simple en carnívoros y animales que se alimentan de concentrados de plantas, y más largo y complejo en omnívoros. Sin embargo, las bajas concentraciones de nutrientes solubles y el alto contenido de carbohidratos estructurales, principalmente celulosa y hemicelulosa, en la dieta de los herbívoros, requieren mecanismos más eficientes para procesar sus alimentos, y sistemas más complejos para la retención y fermentación microbiana del material de las plantas, como en el caso de Alouatta (Finegan & Stevens 2000).
Boca
La cavidad bucal tiene poca capacidad. Sobre el paladar blando se presenta la úvula, de manera rudimentaria en atélidos y cébidos, y muy corta en calitrícidos. Es usual la presencia de surcos medios dorsal y ventral. En el piso de la cavidad bucal un par de pliegues mucosos forman la lamella, en la que desembocan los ductos de Wharton (Varela 2005). La lengua es un órgano muscular en el que se diferencian el vértice, el cuerpo y la raíz (Mayor & López 2012). Todo el dorso de la lengua tiene papilas que varían en número y disposición y pueden ser tanto mecánicas (filiformes) como gustativas (fungiformes, circunvaladas y foliadas) (Varela 2005, Mayor & López 2012).
Las glándulas salivales son comparativamente grandes y están en cantidad variable, pero siempre presentes, la parótida, submandibular y lingual (Varela 2005).
Estómago
El estómago de los primates del nuevo mundo es monocavitario. Se dispone entre el cardias, que comunica con el esófago y está ubicado en el lado izquierdo, y el píloro, que comunica con el duodeno y está situado en el lado derecho. La curvatura mayor del estómago se sitúa caudoventralmente y la curvatura menor, craneodorsalmente. Como en los carnívoros, en el estómago se puede reconocer un fundus poco desarrollado, un cuerpo y una parte pilórica compuesta por el antro y canal pilórico. Toda la mucosa del estómago o gran parte de ella tiene carácter glandular (Mayor & López 2012).
Intestino
El intestino de los primates alcanza de una a tres veces la longitud corporal, siendo considerablemente más corto que el de los herbívoros, donde llega a tener una longitud 25 veces mayor que la longitud corporal (Mayor & López 2012).
El duodeno y el colon están fijados por pliegues peritoneales. Esta disposición, adicional a la presencia de una válvula íleocecal bien desarrollada, contribuyen fisiológicamente al incremento de volumen en la parte posterior del intestino, y la tendencia a acumular materia fecal (Varela 2005).
Intestino delgado
El intestino delgado es largo, especialmente en las especies consumidoras de hojas, pero el mesoduodeno es corto o ausente (Varela 2005). Sus asas se disponen en el espacio comprendido entre el hígado y el estómago (cranealmente) y la entrada de la pelvis (caudalmente). Está formado por tres porciones (Mayor & López 2012):
➢ Duodeno. La porción más proximal del intestino delgado, se extiende entre el píloro y la flexura duodenoyeyunal.
➢ Yeyuno. Es la porción de mayor longitud.
➢ Íleon. Es un segmento corto que se une al ciego por medio del pliegue ileocecal. Su gruesa capa muscular evita el reflujo del contenido intestinal desde el intestino grueso.
Intestino grueso
El intestino grueso se extiende desde el orificio ileal hasta el ano. Sus tres partes son (Varela 2005, Mayor & López 2012):
➢ Ciego. Es la porción más inconstante del intestino grueso. Es un órgano saculado, provisto de bandas tendinosas, que pueden continuar sobre el colon cuando es saculado, pero sin flexura sigmoide; en la parte distal hay una pared especializada provista de tejido linfoide. El volumen cecal suele estar relacionado con la proporción de celulosa del alimento.
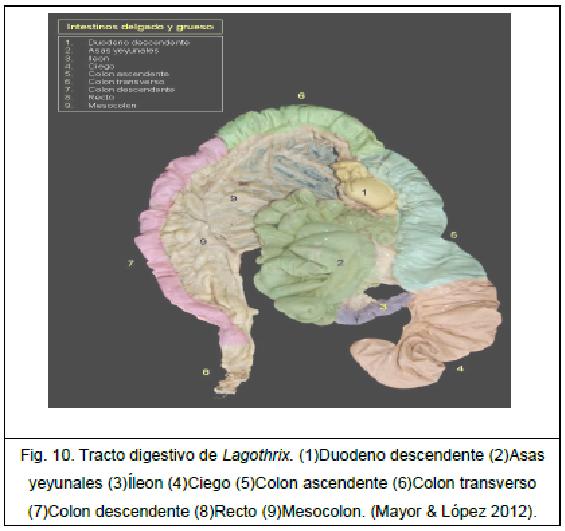
➢ Colon. Está formado por el colon ascendente, dispuesto a la derecha de la cavidad abdominal; el colon transverso, dispuesto transversalmente por detrás del estómago y pasando de derecha a izquierda de la cavidad abdominal; y el colon descendente, que es más largo y llega hasta la entrada de la cavidad pélvica. Tiene una gran capacidad, es largo en calitrícidos, aótidos y en el género Lagothrix, siendo el mesocolon siempre corto. No presentan cisterna del quilo.
➢ Recto. Está situado en la cavidad pélvica y finaliza en el canal anal. Es un órgano tubular relacionado con el colon terminal.
Hígado
El hígado se encuentra protegido por las costillas en la porción intratorácica de la cavidad abdominal. Presenta una cara diafragmática, en contacto con el diafragma, y una cara visceral, en contacto con el estómago. En los primates, los lóbulos hepáticos se encuentran separados por profundas incisuras o fisuras interlobulares (Varela 2005, Mayor & López 2012).
El patrón lobular del hígado, aunque presenta algunas diferencias interespecíficas, es el siguiente (Varela 2005, Mayor & López 2012):
➢ Lóbulo lateral izquierdo
➢ Lóbulo medial izquierdo
➢ Lóbulo cuadrado o central
➢ Lóbulo caudado, en el que se diferencian el proceso papilar y el proceso caudado
➢ Lóbulo medial derecho
➢ Lóbulo medial izquierdo
En los primates del nuevo mundo, las fisuras están especialmente marcadas, lo que permite el deslizamiento de los lóbulos entre sí durante los amplios movimientos de extensión y flexión del tronco. Es importante recordar que estas especies, por su condición de arborícolas, muestran una gran flexibilidad corporal. La vesícula biliar se encuentra entre el lóbulo cuadrado y el lóbulo medial derecho(Mayor & López 2012).
Páncreas
El páncreas tiene un cuerpo relativamente sólido, se encuentra situado de manera transversa en el abdomen. Uno o dos ductos pueden estar presentes, desembocando junto con el ducto biliar en el duodeno (Varela 2005).
Variaciones anatómicas
Cebidae
La lengua de los géneros Cebus y Saimiri es de color negro y presenta papilas mecánicas (filiformes) y papilas gustativas (fungiformes, circunvaladas y foliadas) (Mayor & López 2012).
En la especie Cebus albifrons la longitud total del intestino mide 2.57 veces la longitud corporal, mientras que en el Saimiri sciureus tiene una media de 1.66 veces la longitud corporal. En ambas especies la totalidad de la mucosa del estómago tiene carácter glandular (Mayor & López 2012).

El ciego de los cébidos presenta un mínimo desarrollo. En el caso de esta familia, es importante destacar que en comparación con otros primates del nuevo mundo, suele ingerir vertebrados e invertebrados. El colon es muy sencillo, con un colon ascendente muy corto. El diámetro del colon es similar al del yeyuno (Mayor & López 2012). El sector caudal sacro del recto muestra una dilatación ampular, básicamente una condensación de fibras musculares sobre la pared ventral (Varela 2005).
En el género Cebus, las fisuras hepáticas están especialmente marcadas, lo que permite el deslizamiento de los lóbulos entre sí durante los amplios movimientos de extensión y flexión del tronco. Es importante recordar que esta especie, por su condición de arborícola, muestra una gran flexibilidad corporal (Mayor & López 2012). El lóbulo caudado es de gran tamaño y cubre el riñón derecho (Varela 2005).
Aotidae
En la lengua del mono nocturno se observan papilas mecánicas (filiformes) y gustativas (fungiformes, circunvaladas y foliadas) (Mayor & López 2012). La glándula submandibular es simple (Varela 2005). La mayor parte de la mucosa del estómago tiene carácter glandular (Mayor & López 2012).

En el mono nocturno la longitud total del intestino alcanza aproximadamente 3 veces la longitud corporal del animal. El ciego presenta un cierto desarrollo, hecho posiblemente relacionado con la dieta principalmente frugívora de la especie (Mayor & López 2012).
El colon, aunque sencillo, muestra un tamaño importante. El tramo más próximo al ciego está considerablemente dilatado y forma la ampolla del colon ascendente; la parte restante del colon ascendente se adelgaza para continuarse con el colon transverso. El diámetro de la parte final del colon ascendente, del colon transverso y del colon descendente es similar al del yeyuno (Mayor y López 2012).
Pitheciidae
El sistema digestivo presenta algunas diferencias con respecto al resto de los platirrinos. La morfología dental está bien adaptada a la extracción y masticación de semillas, presentan un intestino grueso más largo que los frugívoros y el lóbulo hepático caudado cubre el riñón derecho (Norconk et al 2002, Varela 2005).
Atelidae
La lengua es pigmentada frecuentemente, en su borde dorsal es rosada o ligeramente azulosa; Ateles geoffroyi grisesens tiene un patrón blanco sobre una base negra. La glándula salival submandibular en los atélidos posee elementos adicionales profundos al milohioides; algunos géneros como Ateles y Lagothrix, exhiben glándulas secundarias bien definidas con ductos propios. La glándula sublingual, únicamente descrita en Ateles, se desarrolla en el piso de la cavidad bucal (Varela 2005).
El estómago en Ateles y Alouatta presenta una incipiente tendencia a la saculación (Varela 2005). En Ateles chamek toda la mucosa del estómago tiene carácter glandular. La longitud total del intestino alcanza 2.54 veces la longitud corporal. Presenta un ciego muy desarrollado. El colon es muy sencillo
y su diámetro es similar al del yeyuno (Mayor & López 2012).

En el género Alouatta, el color de los dientes coincide con el consumo de material vegetal, debido a la presencia de taninos en las especies vegetales consumidas (Pastor-Nieto 2004). En una posición lateral a la raíz de la lengua y ya en la orofaringe se encuentran las tonsilas palatinas (Mayor & López 2012).
El estómago de Alouatta exhibe una gran cantidad de adaptaciones cuando se compara con el de otros atélidos. El saco globular tiene gran capacidad y se estrecha cerca del píloro (Mark & Edwards 2009).Toda la mucosa del estómago tiene carácter glandular (Mayor & López 2012).
Los primates especializados en el consumo de hojas exhiben distintas adaptaciones del intestino, que pueden incluir saculación y expansión del tracto gastrointestinal, dependiendo del sitio de fermentación del alimento. En el género Alouatta se da la fermentación postgástrica, por lo que muestran un ciego desarrollado y colon saculado (Mark & Edwards 2009).

La longitud total del intestino alcanza 3.07 veces la longitud corporal. Este género está considerado como el único vegetariano estricto; posiblemente debido a esta condición este primate es el que presenta el ciego más desarrollado de entre los primates del nuevo mundo (Mayor & López 2012).
El colon ascendente del género Alouatta, aunque sencillo, muestra un tamaño importante. El tramo más próximo al ciego está considerablemente dilatado y forma la ampolla del colon ascendente; la parte restante del colon ascendente se adelgaza para continuarse con el colon transverso (Mayor & López 2012).
Al igual que para el género Alouatta, para el género Lagothrix, en una posición lateral a la raíz de la lengua, y ya en la orofaringe, se encuentran las tonsilas palatinas. Toda la mucosa del estómago tiene carácter glandular. La longitud total del intestino alcanza 2.35 veces la longitud corporal (Mayor & López 2012).

El género Lagothrix presenta un ciego bastante desarrollado, posiblemente debido a su condición de frugívoro y herbívoro. El colon es muy sencillo. El diámetro del colon es similar al del yeyuno (Mayor & López 2012).
Callitrichidae
El paladar está dotado de una papila incisiva y un número variable de pliegues transversos, con una disposición simétrica (Varela 2005). Como en los carnívoros, en el estómago se puede reconocer un fundus poco desarrollado, un cuerpo y una parte pilórica compuesta por el antro y canal pilórico. Toda la mucosa del estómago tiene carácter glandular. La longitud total del intestino alcanza una longitud aproximada de tres veces la longitud corporal. El ciego de los calitrícidos presenta un cierto desarrollo, hecho posiblemente relacionado con la dieta omnívora de la familia y con la proporción de celulosa del alimento. El colon es muy sencillo (Mayor & López 2012).

Requerimientos nutricionales específicos
Vitamina D
Se activa una vez que llega al hígado. Tiene gran importancia en el metabolismo del calcio, principalmente para la síntesis de tejido óseo (Gázquez 1991). Dentro de primates del nuevo mundo, la vitamina D3 ha mostrado ser biológicamente más activa que la vitamina D2 en la absorción de calcio o la prevención y reversión de la enfermedad metabólica ósea. Esta variación ha sido observada en Cebus albifrons, Saimiri sciureus, Saguinus oedipus, S. mystax y S. nigricollis (Hunt et al 1969). Se ha propuesto que los tamarinos y marmosetas requieren cantidades mayores de vitamina D3 que otras especies de primates del nuevo mundo, debido a la resistencia de los órganos receptores a la forma activa de la vitamina (Crissey et al 2003).
 |
| Tabla de los principales requerimientos nutricionales en primates del nuevo mundo. |
Como puede observarse, el género Alouatta requiere una mayor cantidad de fibra que el resto de los primates debido a las adaptaciones anatomofisiológicas que lo caracterizan como folívoro (Edwards 2009).
Principales patologías nutricionales
Diversas enfermedades metabólicas están relacionadas con un mal balanceo de la dieta de los primates.
Enfermedad metabólica ósea
El calcio y el fósforo son los principales componentes minerales del tejido óseo. Sin embargo para mantener la estabilidad de su estructura, se requieren niveles adecuados de cobre, yodo y zinc. Por lo tanto, estos oligoelementos no deben ser olvidados, tanto al formular raciones para animales en crecimiento, gestación o lactancia, como al momento de establecer diagnósticos diferenciales de causas de patologías óseas (Unger & Chiappe 2008).
Los primates del nuevo mundo no pueden utilizar la vitamina D2, por lo que requieren la suplementación dietética de colecalciferol. La enfermedad metabólica ósea en estos primates puede producirse por la deficiencia de vitamina D3 en la dieta; y se ha propuesto que una dieta no balanceada en calcio y fósforo, por la utilización excesiva de frutas en la dieta, puede influir (Baskin 2008).
 |
| Fractura humeral distal en Alouatta seniculus. Hueso hipodenso |
Animales con debilidad, desmineralización ósea, fracturas múltiples, cifosis y huesos encorvados, son el resultado de dietas mal balanceadas en minerales. Microscópicamente, la alta actividad osteoclástica conduce a desmineralización ósea, hipertrofia paratiroidea y se presenta glomerulonefritis membranosa y nefritis intersticial crónica (Hunt et al 1969, Crissey et al 2003). A largo plazo provoca hiperparatiroidismo nutricional secundario, lo cual resulta en resorción ósea y reemplazo fibroso (Baskin 2008). El raquitismo es un desorden comúnmente observado en primates no humanos mantenidos en cautiverio (Fowler & Miller 2012).
Escorbuto
Se llama escorbuto a la enfermedad causada por la deficiencia de vitamina C en la dieta. Clínicamente se han reportado cefalohematomas en monos ardilla y problemas de anemia en este y otros primates, debido a que no poseen la enzima L-gulonolactona oxidasa, necesaria para formar el ácido ascórbico (Cudós & Diángelo 1996, Edwards 2009, Fowler & Miller 2012).
Los animales afectados responden de manera favorable a la administración de vitamina C a dosis de 7.5-10mg/Kg, aunque las necesidades para cada especie no han sido completamente establecidas (Fowler & Miller 2012). El principal defecto causado por la deficiencia de vitamina C, es la incapacidad para formar colágeno normal. Esto se manifiesta por hemorragias subperiósticas, fracturas epifisarias y osificación anormal de los huesos, en los que pueede observarse una amplia zona de cartílago calcificado, pero no osificado (Crissey et al 2003, Baskin 2008).
Obesidad
Dentro de los factores que predisponen a la obesidad en animales en cautiverio, se encuentran las dietas hipercalóricas, falta de actividad física, genética, y contracepción en hembras. La obesidad incrementa la presión sobre las articulaciones y la enfermedad degenerativa del músculo esquelético en animales de edad avanzada, además de predisponer a neoplasia y desórdenes metabólicos como el desarrollo de diabetes o lipidosis hepática (Fowler & Miller 2012).
Diabetes
La diabetes mellitus, caracterizada por degeneración severa difusa de los islotes de Langerhans ha sido diagnosticada en Leontopithecus chrysomelas y Leontopitecus chrysopygus. Los cambios grasos del hígado, arteriosclerosis de las capas media e interna de la aorta, nefritis intersticial bilateral crónica e hidronefrosis fueron también observadas en tamarinos (Fowler & Miller 2012).
Síndrome de emaciación de la marmoseta
El síndrome de emaciación de la marmoseta es una enfermedad nutricional dada por un desequilibrio protéico - calórico. Las concentraciones de energía y proteína en la dieta son muy importantes para prevenirla y deben suministrarse conforme a los requerimientos de la especie (Crissey et al 2003).
Anemia en Aotus
A pesar de ser de etiología aún desconocida, responde favorablemente a la administración parenteral de vitamina E y selenio, incluso cuando los niveles en la dieta y el suero parecen adecuados. Algunos de los 11 cariotipos son susceptibles, mientras que otros no (Baskin 2008).
Las enteropatías podrían favorecer la malabsorción de vitamina E. Clínicamente se observa anemia hemolítica, palidez de membranas mucosas y otros tejidos, ictericia, petequias en sistema nervioso central, glóbulos rojos nucleados circulantes, necrosis centrolobulillar hepática, hemosiderosis, microinfartos cerebrales, edema, ascitis, cardiomegalia y necrosis muscular (Baskin 2008).
Enfermedad periodontal
Esta enfermedad se caracteriza por la inflamación de las encías o gingivitis, con presencia de sarro alrededor de los dientes. Sus principales complicaciones incluyen la pérdida de piezas dentales, infecciones sistémicas que viajan vía hematógena o linfática a partir del drenaje de la cavidad bucal y anorexia por dolor (Fowler & Miller 2012). Los animales que consumen alimentos blandos en exceso son más susceptibles a padecer esta enfermedad (Crissey et al 2003).
Técnicas de Alimentación
La mayoría de las especies de primates son omnívoras. Sin embargo, no debe ser ofrecida la misma fórmula alimenticia para todas las especies (Edwards 2009). Los alimentos comerciales para perros fueron inicialmente usados para suplementar las dietas de los primates en cautiverio, y en muchos zoológicos todavía son usados estos alimentos. Algunos primates son alimentados con productos de origen animal, tales como carne, corazón o pollo, adicionando leche en polvo, queso, yogurt, miel, huevo y harinas (Crissey et al 2003, Gómez et al 2006).
En la actualidad, muchos zoológicos centran su alimentación en pellets específicos para primates, suplementando con los ingredientes mencionados. Debido a la naturaleza propia de los primates del nuevo mundo y su metabolismo, la cantidad total de alimento que será ofrecido debe dividirse en dos o más porciones (Pastor-Nieto 2004, Edwards 2009).
Los monos aulladores son considerados como los primates más difíciles de mantener en cautiverio debido a la alta especialización de su dieta en vida libre, la cual contiene un 90% más de fibra en comparación con los alimentos que se ofrecen habitualmente en los zoológicos. Aunado a esto, la mayoría de los ejemplares del género Alouatta ingresan a cautiverio en pobres condiciones de salud, sobreviviendo pocos meses, debido a una dieta nutricionalmente pobre en hojas, tallos, brotes y flores. La adaptación anatomofisiológica que permite a los primates folívoros utilizar eficientemente las plantas como principal fuente de nutrientes, limita su adaptabilidad a las dietas comerciales disponibles para la mayoría de primates .(Pastor-Nieto 2004, Edwards 2009, Anaya et al 2010).
Algunas condiciones pueden ser evitadas con un correcto manejo nutricional. La restricción alimenticia ha resultado en baja incidencia de osteoartritis, ya que reduce el peso corporal, y por lo tanto el desgaste articular. La restricción de proteina y fosfato incrementa la media de supervivencia en animales con enfermedad renal (Fowler & Miller 2012).
Consideraciones generales para la alimentación en cautiverio
Para poder dar a los animales un manejo nutricional acorde a sus necesidades, es importante considerar:
➢ Edad. Existen varias características anatómicas para determinar indirectamente la edad de los animales, como el peso, la presencia y color de los dientes, presencia de testículos en escroto, pigmentación de la piel en Ateles (Pastor-Nieto 2004, Varela 2005).
➢ Estado fisiológico. Los requerimientos aumentan durante la gestación y lactancia.
➢ Examen clínico general. Este debe incluir el peso y la observación de signos vitales, letargo e hipotermia. Cuando los animales provienen de decomisos, es común observarlos mal nutridos, deshidratados, hipotérmicos, hipoglucémicos y enfermos. Se debe realizar biometría hematica para descartar infecciones, anemia, y parásitos sanguíneos, análisis coproparasitoscópicos para identificar parásitos adquiridos en vida libre o en cautiverio, coprocultivo para determinar si son portadores de enterobacterias zoonóticas (Pastor-Nieto 2004). Todos estos factores influirán en la dieta a ser ofrecida al ejemplar.
➢ Comportamiento. Muchos especimenes están improntados o acostumbrados al contacto con seres humanos. Otros se encuentran seriamente estresados y en consecuencia inmunosuprimidos. Resulta necesario proveerles de los elementos necesarios para enriquecer su medio ambiente y cuando sea posible, permitir una adecuada integración social con otros de su especie (Pastor-Nieto 2004).
➢ Estacionalidad. El consumo de alimento puede variar de acuerdo a la estación del año,como un comportamiento normal de la especie. Esto puede influir en la masa corporal del individuo, por lo que debe ser considerada en la formulación de la dieta (Crissey et al 2003).
Lactantes
Existen varias razones para la crianza artificial de primates en cautiverio. Las principales incluyen el tráfico ilegal, la muerte de la madre o la agresión a la cría por parte del grupo. La mayoría de los lactantes que son víctimas del tráfico ilegal, padecen hipotermia, hipoglucemia y deshidratación, para lo cual es necesario administrar fluidos y dar terapia de sostén, que puede ser por vía oral o parenteral, dependiendo de la severidad del caso. En la mayoría de los casos estas soluciones se tienen que suministrar tibias y de manera forzada (NRC 2003, Pastor-Nieto 2004).
 |
| Mono araña. Parque Zoológico Nacional la Aurora, Guatemala. Foto: Andrea Castañeda. |
Una vez estabilizado se debe alimentar al lactante con fórmulas específicas para primates, o en su defecto con fórmulas humanas maternizadas, de preferencia suplementadas con hierro. Si el reflejo de succión está presente, se pueden utilizar biberones para primates o para la alimentación de crías huérfanas de especies domésticas, o jeringas adaptadas al chupón y verificar el flujo de este. Por lo general no requieren estimulación perineal para fomentar la micción y defecación (Pastor-Nieto 2004).
Infantes
Después de un periodo aproximado de cuatro a seis semanas de alimentar a la cría con fórmulas lácteas, se puede empezar a ofrecer alimentos sólidos o en puré. La inclusión de alimentos comerciales para niños, elaborados a base de frutas y verduras, tiene grandes ventajas, ya que son nutritivos e higiénicos y vienen adicionados con vitaminas y minerales (Pastor-Nieto 2004).
Una mejor opción es ofrecer purés naturales de frutas y pedacitos de verduras cocidas o bien desinfectadas. Inicialmente, es muy posible que los infantes no manifiesten interés hacia estos alimentos. Se debe seguir ofreciendo fórmula láctea maternizada durante este periodo, permitiendo la adaptación del animal al consumo de los nuevos ingredientes (Pastor-Nieto 2004).
 |
| Mono aullador. Parque Zoológico Nacional la Aurora, Guatemala. |
Juveniles y sub-adultos
En esta etapa, es importante diferenciar el manejo nutricional de cada especie, dependiendo de su ecología conductual. Los miembros del género Ateles son preponderantemente frugívoros, mientras que los del género Alouatta son esencialmente folívoros, y el resto omnívoros, diferencia que marca el tipo de manejo que se debe dar a cada uno (Pastor-Nieto 2004, Richard 2004).
Se estima que los primates del género Alouatta obtienen del 26 al 36% de sus requerimientos diarios de energía a través de los ácidos grasos volátiles, producto de la fermentación microbiana que sufren hojas y frutos en el ciego y colon. La energía obtenida a través de este proceso depende de la cantidad y calidad de forraje que ellos consuman (Anaya et al 2010).
Lo anterior debe considerarse responsablemente, ya que los tractos gastrointestinales están adaptados evolutivamente a estas dietas. En cautiverio se recomienda dividir la ración en dos partes, además de proveer agua ad libitum (Pastor-Nieto 2004, Anaya et al 2010).
Aquellas instituciones que han tenido mayor éxito en el mantenimiento de folívoros en cautiverio, son las que han cubierto sus necesidades especializadas. Existen algunas formas de incrementar la cantidad de fibra en la dieta de estas especies. La más difundida y fácil de implementar es el reemplazo de concentrados especiales para primates, que tienen un bajo nivel de fibra (10-19% FDN, 5-7.5% FDA) por uno con una mayor cantidad de fibra (25% FDN, 15% FDA), ya que los primeros fueron diseñados para primates omnívoros (Edwards 2009).
Otra opción es la reducción o eliminación de frutas disponibles en el mercado, reemplazándolas por hojas verdes y material vegetal alto en fibra. Este método se utiliza con frecuencia en combinación con la incorporación de concentrados especiales para primates, altos en fibra (Edwards 2009).
El cultivo de plantas específicas para suplementar a estas especies es otra opción. Es una práctica que no se ha distribuido en gran medida, debido principalmente a las condiciones locales que impiden su crecimiento o a factores económicos. Los tipos de plantas cultivables pueden incluir desde especies nativas hasta especies exóticas de rápido crecimiento, prefiriéndose aquellas que ellos consumirían en la naturaleza (Edwards 2009).
La composición nutricional de las plantas ofrecidas en cautiverio debe ser determinada, así como el impacto estacional en variabilidad y la fertilización de las mismas. Se deben vigilar las porciones consumidas por los animales y la salud de los mismos, ya que se han reportado muertes atribuibles a la alimentación con ciertas plantas en animales en cautiverio, debidas tanto a las defensas desarrolladas por las plantas, como a la falta de habituación por parte de los individuos en cautiverio, a consumir estas especies (Edwards 2009).
Se sugiere que una aproximación ideal a la alimentación de primates folívoros en cautiverio, debe considerar la combinación de (Edwards 2009):
➢ Uso de alimentos específicos para primates, que sean altos en fibra.
➢ Reducción o eliminación de frutas e incremento de hojas verdes.
➢ Si se da forraje natural, dar las plantas apropiadas, que no sean tóxicas y que simulen la composición y proporción de la dieta natural de la especie en cuestión.
➢ Distribuir estos alimentos en pequeñas cantidades durante todo el día.
El consumo de materia vegetal es crucial para estimular el buen tránsito intestinal en Alouatta, además de fomentar el hábito natural de forrajeo. Por lo anterior, es importante adaptar a este género al consumo de especies vegetales de la región que puedan ser consumidas por ellos (Pastor-Nieto 2004).
Una dieta balanceada nutricionalmente es importante para el mantenimiento de la salud bucal. Se deben ofrecer periódicamente alimentos crujientes o duros para ayudar en el mantenimiento de la higiene dental (Crissey et al 2003).
Con respecto a las formas de administración de alimentos, se sugiere tener algunas consideraciones:
➢ Debido a que los primates del nuevo mundo son de hábitos arborícolas, estos animales poseen bajas defensas para patógenos terrestres, por lo que se sugiere adaptar los comederos en alto, sujetos a la malla de la jaula o en una plataforma, y no dar el alimento en el piso, para que no se contamine con heces fecales y orina (Pastor-Nieto 2004).
➢ Es preferible revolver los ingredientes de la dieta, ya que los animales seleccionan el alimento que desean consumir, además el uso de saborizantes puede hacer más palatables ingredientes de alto valor nutricional que los animales no consuman regularmente.
➢ Se debe considerar el balance correcto de la dieta acorde al periodo fisiológico en el que se encuentra el animal, tal como el crecimiento, gestación o lactancia, durante los cuales los niveles de energía, proteina y los de algunos minerales, pueden incrementar de manera sustancial (Serio-Silva et al 1999).
➢ Deben tomarse precauciones al formular las dietas de animales gestantes, ya que dietas altas en grasas han sido relacionadas con una reducción de ácidos grasos n-3 en el plasma y con apoptosis hepática fetal en primates (Grant et al 2011).
Algunos estudios experimentales han demostrado la influencia que tiene una dieta adecuada durante la gestación, en la fertilidad de las crías, cuando estas alcanzan su madurez. Se observó que las hembras provenientes de animales desnutridos durante la gestación, tendían a tener crías más pequeñas. Asimismo, se vio afectada la edad a la cual gestaban por primera vez, y la mortalidad pre y perinatal (Gardner et al 2009).
Alimentos usados en cautiverio
Cebidae
Para los cébidos, la dieta debe ser variada y omnívora. Los principales alimentos utilizados en cautiverio son huevo crudo o hervido, carne, semillas, frutas varias, principalmente papaya, sandía, manzana y banano, y vegetales, como brócoli y zanahoria, suplementado con alimentos comerciales para primates omnívoros (Agudelo et al 2005).
Aotidae
En colonias mantenidas en cautiverio se han utilizado dietas compuestas por frutas de temporada, entre estas plátano, guayaba, papaya, piña, mango, acompañada de alimentos comerciales para primates que contienen entre 15 y 25% de proteína, 5% de grasas, vitaminas y minerales (Umaña et al 1984).
Pitheciidae
Su alimentación en cautiverio se basa en alimento comercial para primates, frutas, vegetales y suplementos vitamínicos (Norconk et al 2002).
Atelidae
La dieta de Ateles consiste generalmente de frutas de temporada, dependiendo de su costo, tales como plátano, papaya, naranja, piña, manzana, pera; y algunos vegetales, aunque en menor proporción que las frutas, como lechuga, tomate, zanahoria, cilantro, apio. También se le incluye
alimento comercial para primates del nuevo mundo de Purina o Mazuri, por ser un alimento balanceado específicamente diseñado para esta especie. Sin embargo su venta comercial no es común en los países latinoamericanos, por lo que en algunos países aún se utiliza alimento comercial para perros, remojado en jugos de frutas (Pastor-Nieto 2004).
La dieta de Alouatta debe consistir preponderantemente de verduras bien desinfectadas, como cilantro, apio, espinacas, alfalfa, y algunas frutas en trozos (Pastor-Nieto 2004, Anaya et al 2010), como mango, piña y papaya (Dierenfeld & Graffam 1996). Si es factible se pueden ofrecer otras especies vegetales, no tóxicas, para fomentar la conducta natural de forrajeo (Anaya et al 2010).
La suplementación esporádica con proteína de origen animal se recomienda, ya que los monos aulladores parecen obtener un buen balance de aminoácidos al consumir brotes tiernos de una amplia variedad de especies vegetales en vida libre. Para tal efecto se utiliza alimento comercial para bebé a base de res o pollo, o alimento enlatado para perro, bajo en grasa. También se utilizan alimentos específicos para primates o folívoros, cereales y algunos derivados lácteos (Pastor-Nieto 2004, Anaya et al 2010).
Tanto para Ateles como para Alouatta, se han utilizado alimentos comerciales de uso humano, a manera de aportar proteínas, carbohidratos, vitaminas, minerales y oligoelementos (Pastor-Nieto 2004).
Para la alimentación del género Lagothrix en cautiverio, se han utilizado ingredientes como banano, papaya, mango, lechuga, frijol pinto, mazorca dulce, concentrado para omnívoros, huevo cocido y concentrado para perro (Dierenfeld & Graffam 1996).
En el caso de los lactantes de Alouatta y Ateles, se han utilizado de manera exitosa, fórmulas maternizadas humanas enriquecidas con polivitamínicos pediátricos, disueltos en agua hervida o purificada (Pastor-Nieto 2004).
Callitrichidae.
Generalmente, las dietas ofrecidas a los calitrícidos mantenidos en cautiverio se componen de frutas (higo, manzana, banana, mango, papaya, guayaba), vegetales (brócoli), insectos, huevo, pollo y alimentos comerciales para primates (Crissey et al 2003, Gómez et al 2006). En algunos zoológicos, se han incluido los exudados vegetales y goma arábiga (NRC 2003).
Los principales ingredientes utilizados para la formulación de raciones de Saguinus spp. mantenidos en cautiverio son banano, uva, papaya, piña, mango, frijol, lechuga, espinaca, acelga, concentrado para omnívoros y mezclas de carne (Dierenfeld & Graffam 1996).
Referencias bibliográficas
- Agudelo, N., K. Montoya, L.M. Gutiérrez, S. Osorio & D.I. Balvín. 2005. Establecimiento de un Protocolo de Liberación de Cebus capucinus en el Centro de Atención y Valoración de Especies de Fauna Silvestre (CAV) Barbosa – Antioquia. Resultados Preliminares. Libro de Resúmenes del I Congreso Internacional de Medicina y Aprovechamiento de Fauna Silvestre Neotropical. Facultad de Medicina Veterinaria y Zootecnia. Universidad Nacional de Colombia. Bogotá, Colombia. 85-97.
- Anaya, M., P. Cifuentes, A. Ducoing, M. Sánchez & C. Gutiérrez. 2010. Determinación de nutrientes en la dieta ofrecida a monos aulladores (Alouatta palliata y A. pigra.) en el Zoológico de Chapultepec. REDVET. Rev. electrón. vet. 11: 1-10.
- Barbosa, C. 2008. Ecologia alimentar, padrão de atividade e uso de espaço por Callicebus nigrifrons (Primates: Pitheciidae). Universidade Estadual de Campinas . Instituto de Biologia. (Consultado: 11 febrero 2012, http://www.universia.net).
- Baskin, G.B. 2008. Pathology of Nonhuman Primates. WRPRC Primate Information Network. (Consultado: 13 febrero 2012, http://pin.primate.wisc.edu/research/vet/pola6-99.html).
- Chatterjee, H.J., S.Y. Ho, I. Barnes & C. Groves. 2009. Estimating de phylogeny and divergence times of primates using a supermatrix approach. BMC Evolutionary Biology 9: 1-19.
- Crissey, S.D., M. Gore, B.A. Lintzenich & K. Slifka. 2003. Callitrichids: Nutrition and Dietary Husbandry. Nutrition Advisory Group Handbook.
- Crissey, S.D., M. Maslanka & D.E. Ullrey. 2002. Assessment of Nutritional Status of Captive and Free-Ranging Animals. Nutrition Advisory Group Handbook.
- Cudós, A. & A.M. Diángelo. 2007. Percepción materna del estado nutricional y de la ingesta alimentaria de niños preescolares. Tesis de la Carrera de Licenciatura en Nutrición. Centro Regional Rosario. Universidad de Concepción del Uruguay.
- Dierenfeld, E.S. & W.S. Graffam. 1996. Manual de Nutrición y Dietas para Animales Silvestres en Cautiverio (ejemplos para Animales de América Latina)
- Edwards, M.S. 2009. Primate Nutrition: Novel Feeding Options for Folivorous Species. (Consultado: 7 febrero 2012, http://www.colyerinstitute.org).
- Finegan, E.J. & E. Stevens. 2000. The Digestive System of Vertebrates Website. (Consultado: 11 febrero 2012, http://www.cnsweb.org/digestvertebrates/AAAEdStevensINTRODUCTION.html).
- Fowler, M.E. & Miller, R.E. 2012. Fowler’s Zoo and Wild Animal Medicine. Current Therapy. Saunders, Elsevier Inc. St Louis Missouri, U.S.A.
- Gardner, D.S., S.E. Ozanne & K.D. Sinclair. 2009. Effect of the early-life nutritional environment on fecundity and fertility of mammals. Phil. Trans. R. Soc. B 364: 3419-3427.
- Gázquez,A. 1991. Patología Veterinaria. Primera edición. Ed. Interamericana McGraw-Hill. Madrid, España.
- Gómez S., I. Lozano, A. Jaramillo & A. Arias. 2006. Establecimiento y Evaluación de una Dieta para Monos Tití Gris (Saguinus leucopus, Günter, 1876) y Estudio del Comportamiento Alimentario en Cautiverio en la Fundación Zoológico Santacruz (Colombia - Sur América). Mem. Conf. Interna Med. Aprovech. Fauna Silv. Exót. Conv. 2: 9-18.
- Goyoaga, C. 2005. Estudio de factores no nutritivos en Vicia faba I.: Influencia de la germinación sobre su valor nutritivo. Tesis Doctoral. Universidad Complutense de Madrid.
- Grant, W.F., M.B. Gillingham, A.K. Batra, N.M. Fewkes, S.M. Comstock, D. Takahashi. T.P. Braun, K.L. Grove, J.E. Friedman & D.L. Marks. Maternal High Fat Diet Is Associated with Decreased Plasma n-3 Fatty Acids and Fetal Hepatic Apoptosis in Nonhuman Primates. 2011. 6:1-14.
- Hunt, R.D., F.G. García & D.M. Hegsted. 1969. Hypervitaminosis D in New World Monkeys. The American Journal of Clinical Nutrition. 22: 358-366.
- Mayor, P & C. López. 2012. Atlas de Anatomía de Especies Silvestres de la Amazonia Peruana. (Consultado: 10 febrero 2012, http://atlasanatomiaamazonia.uab.cat).
- National Research Council (NRC). 2003.Nutrient Requirements of Nonhuman Primates. Washington, DC: National Academy Press.
- Norconk, M.A., O.T. Oftedal, M.L. Power, M. Jakubasz & A. Savage. 2002. Digesta Passage and Fiber Digestibility in Captive White-Faced Sakis (Pithecia pithecia). Am. J. of Primatol. 58: 23-34.
- Pastor-Nieto, R. 2004. Manejo en Cautiverio de Alouatta y Ateles. WRPRC Primate Information Network.
- Pinto, L.P. 2008. Ecologia alimentar do cuxiu-de-nariz-vermelho Chiropotes albinasus (Primates: Pitheciidae) na Floresta Nacional do Tapajos, Para. Universidade Estadual de Campinas. Instituto de Biologia. (Consultado: 11 febrero 2012, http://www.universia.net).
- Richard, T. 2004. Historia Natural de los Primates Colombianos. Ed. Universidad Nacional de Colombia. Bogotá, Colombia.
- Riveros, M. & C. Ferreira. 2001. Comportamiento alimentario de una tropa de Pithecia pithecia (mono viudo cara blanca), en una isla del embalse de Guri, Estado Bolívar. ECOTROPICOS 14:57-64.
- Serio-Silva, J.C., L.T. Hernández-Salazar & V. Rico-Gray. 1999. Nutritional Composition of the Diet of Alouatta palliata mexicana Females in Different Reproductive States. Zoo Biology 18: 507-513.
- Schneider, H. 2000. The Current Status of the New World Monkey Phylogeny. An. Acad. Bras. Ci. 72: 165-172.
- Umaña, J.A., J.Ramírez, C.A. Espinal & E. Sabogal. 1984. Primates no Humanos para Investigación Biomédica.
- Establecimiento, Adaptación y Mantenimiento de Aotus lemurinus griseimembra. Bol Of Sanit Panam. 97: 44 53.
- Unger, M. & M.A. Chiappe. 2008. Importancia fisiológica de los microminerales en el metabolismo óseo. REDVET. Rev. electrón. vet. 9: 1-17.
- Valencia, F.E. & M.O. Román. 2004. La fibra dietaria como alimento funcional. Vitae 11:12-17.
- Varela, N. 2005. Consideraciones Anatómicas de Importancia Clínica en los Primates Neotropicales. Revista de la Asociación de Veterinarios de Vida Silvestre 1: 15-27.